

Zakażenia wirusem grypy A (IAV) są powszechne u świń na świecie (Brown, 2000). Zakażenie wiąże się z wystąpieniem ostrych objawów ze strony układu oddechowego, którym często towarzyszą zakażenia bakteryjne co prowadzi do padnięć u młodych prosiąt i zmniejszonych przyrostów w okresie tuczu lub ronienia loch. Tak więc skutki ekonomiczne zakażenie IAV mogą być znaczne (Rajao et al., 2014; Vincent et al., 2014; Vincent et al., 2014).
Świńskie wirusy grypy A należą do rodziny Orthomyxoviridae; posiadają genom zbudowany z pojedynczej nici RNA, złożony z 8 segmentów. Podtypy są identyfikowane na podstawie charakterystyki genetycznej i antygenowej glikoprotein powierzchniowych, hemaglutyniny (HA) i neuraminidazy (NA) (Cheung et al., 2007).

Wirus grypy łączy się z receptorami na powierzchni komórek układu oddechowego, którymi są glikany z kwasem sialowym. Na skutek ekspresji w górnych drogach oddechowych świni receptorów z kasem sialowym w pozycji α-2-6 lub α-2-3, zwierzęta te są wrażliwe na zakażenie szczepami pochodzenia ludzkiego i ptasiego (Byrd-Leotis et al., 2014; Trebbien et al., 2011; Walther et al., 2013). Dlatego też koinfekcje komórek świń szczepami ptasimi i ludzkimi umożliwia reasortację (wymianę segmentów genomów) między nimi (Hass et al., 2011; Ma et al., 2009; Van Poucke et al., 2010; Zell et al., 2013). Reasortacja może być powodem trudnych do przewidzenia zmian fenotypowych wpływających na tropizm wirusa, zjadliwość, strukturę antygenową, odmiennych od szczepów macierzystych.
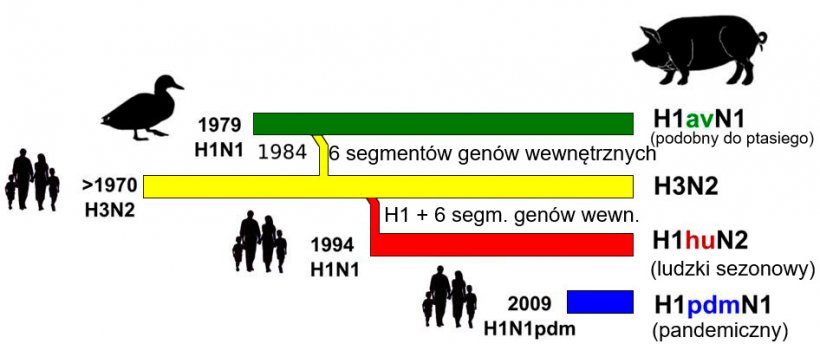
Rycina 1. Historia i pochodzenie wirusów grypy A (IAV) u świń w Europie. Należy podkreślić, że żaden z obecnie krążących u świń szczepów nie pochodzi od tego gatunku zwierząt.
Na rycinie 1 przedstawiono pochodzenie i historię reasortacji szczepów IAV krążących obecnie w populacji świń w Europie. Do 2009 roku w Europie krążyły trzy podtypy IAV, H1N1, H1N2 i H3N2. Najstarszy z podtypów H1N1 został wprowadzony do populacji świń w końcu lat 1970-tych z populacji ptaków, który zastąpił „klasyczny” ludzki wariant z 1918 roku. Ponieważ współczesny podtyp H1N1 pochodzi od ptaków jest określany jako (av)H1N1 (Brown, 2013; Simon et al., 2014). W 1984 roku doszło do reasortacji z wirusem H3N2 wywołującego ludzką grypę sezonową. W efekcie powstał reasortant posiadający 6 wewnętrznych segmentów pochodzenia ptasiego oraz HA (H3) i NA (N2) pochodzenia ludzkiego virus (Kong et al., 2015). Ten wariant wirusa rozprzestrzenił się w kilku krajach europejskich. Kolejny reasortant powstał w 1994 roku w wyniku wprowadzenia do H3N2 segmentu genu HA pochodzącego z innego wirusa ludzkiej grypy sezonowej H1N1. W wyniku tego powstał wirus H1N2. Ten podtyp H1 jest określany jako ludzki (human-like, hu) H1N2 (Brown, 1998). Te linie genetyczne były stabilne i krążyły w Europie z wyjątkiem niektórych obszarów (Brown, 2013; Kuntz-Simon et al., 2009). Wirusy H1N1av były odpowiedzialne za większość zakażeń u świń w Europie (Brown, 2000).
W 2009 roku pojawił się kolejny wirus: ludzki szczep grypy pandemicznej H1N1 (H1N1pdm). Wirus ten jest wynikiem wielokrotnej reasortacji i zawiera amerykańskie segmenty pochodzenia ptasiego (PB2, PA), ludzkie (PB1), klasyczne świńskie (HA, NP, NS), i europejskie świńskie (NA, NP) (Neumann et al., 2009). Ze względu na częściowo świńskie pochodzenie wirusa grypy pandemicznej, infekcje u ludzi zaczęto nazywać „świńską grypą”. Świnie są bardzo wrażliwe na zakażenie tym wariantem i w wielu krajach zaobserwowano zakażania świń przez człowieka. Od tego czasu H1N1pdm rozprzestrzenia się w populacji świń niezależnie od zakażeń u ludzi (Brookes et al., 2010).
Wraz z pojawieniem się tak zwanej “świńskiej grypy” w Europie przeprowadzono program monitoringu grypy świń w latach 2013-2015, projekt ESNIP (Watson et al., 2015). Wyniki pokazały, że H1N1pdm jest obecny w kilku krajach Europy a szczególnie często występuje w Wielkiej Brytanii i Irlandii. Obecność H1N1pdm pochodzenia ludzkiego istotnie zmieniło równowagę epidemiologiczną i doprowadziło do kolejnych reasortacji. W Niemczech pojawiły się reasortanty H i N w wyniku, których powstał wirus H1pdmN2 krążący dotychczas (Starick et al., 2012; Watson et al., 2015). Białko HA tych reasortantów uległo istotnym zmianom i jest antygenowo różne od ludzkiego H1pdm (Harder et al., 2013). Opisano również inne reasoranty wirusa grypy (H1huN1av, H1dpmN1av, H3N1pdm) jednak nie wykryto ich rozprzestrzeniania się w populacji (Watson et al., 2015). Ponadto opisano reasortanty, które wymieniły wszystkie segmenty genów wewnętrznych z wirusami H1N1pdm (Kong et al., 2015; Watson et al., 2015). W USA stwierdzono świńskie szczepy H3N2 lub H1N1 zawierające segment genu M pochodzący z H1N1pdm, które łatwo przenosiły się na człowieka w czasie targów hodowlanych (Schicker et al. 2016). Takie warianty zaobserwowano również w Europie, jednak nie u człowieka. Prewalencja H1N1pdm jest różna w różnych krajach Europy (rycina 2). Inaczej niż na Wyspach Brytyjskich i Irlandii, na kontynencie europejskim H1N1av stale dominuje nad H3N2 i H1huN2 (rycina 2). Podtyp H3N2 nie został stwierdzony w Danii, Polsce i UK, w we Francji stwierdzono jedynie sporadyczne przypadki (Watson et al. 2015).
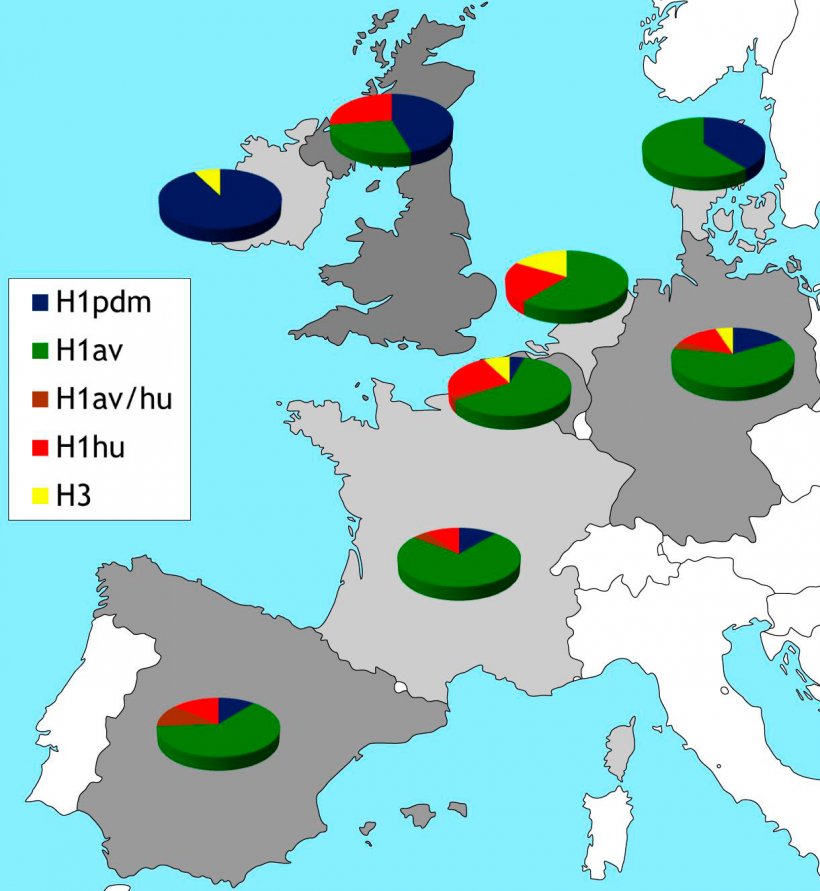
Rycina 2. Geograficzne rozprzestrzenienie podtypów HA IAV krążących obecnie w Europie. Informacje oparte na danych literaturowych i własnym monitoringu biernym przeprowadzonym w roku 2015.
Po zakończeniu projektu ESNIP3 w 2015 roku przeprowadzono monitoring bierny w kilku krajach środkowej i zachodniej Europy. Wstępne dane zgromadzone w pierwszym roku badań wykazały wzrost występowania H1N1pdm i reasortantów tego wirusa. Ponadto stwierdzono wzrost koinfekcji i reasortacji między trzema kocyrkulującymi europejskimi świńskimi wariantami H1. Nie wiadomo czy ten wzrost ma związek z wdrożeniem czulszych narzędzi diagnostycznych w naszym laboratorium: wykrywanie IAV i charakterystyka podtypów HA (H1av, H1pdm, H3) i NA (N1av, N1pdm, N2) jest wykonywana w trzech realckach RT-qPCR. Zwiększona czułość tej metody pozwala na bezpośrednią analizę próbek terenowych (szczególnie wymazów z nosa) bez konieczności izolacji wirusa (Henritzi et al. 2016). Badania te będą kontynuowane do końca 2017 roku.
Ciągły monitoring IAV u świń w Europie jest niezbędny do analizy ewolucji wirusa. Szczepy świńskie mogą odgrywać istotną rolę w tworzeniu nowych szczepów o potencjale pandemiczym jak miało to miejsce w pojawieniu się wirusa H1N1 w 2009 roku. Analiza dynamicznych zmian wśród szczepów IAV i monitoring fenotypów o potencjale zoonotycznym stanowi centralny element polityki OneHealth.
Ostatnie badania nad IAV w laboratorium autorów były finansowane w ramach grantu otrzymanego od IDT, Dessau, Niemcy. Autorzy dziękują Aline Maksimov i Carl Sell za cena pomoc techniczną.
